A Cross-Cultural Analysis of Brazilian, Russian, and American Families

Journal: Social Evolution & History. Volume 24, Number 1 / March 2025
DOI: https://doi.org/10.30884/seh/2025.01.08
Olga Semenova, Institute of Ethnology and Anthropology, RAS, Moscow, Russia
Aurelio José Figueredo, The University of Arizona, Tucson, USA
Rachel Zambrano, The University of Arizona, Tucson, USA
Rosana Suemi Tokumaru, Universidade Federal do Espírito Santo, Brazil
Renata Pereira Defelipe, Universidade Presbiteriana Mackenzie, São Paulo, Brazil
Tania Kiehl Lucci, Universidade de São Paulo, São Paulo, Brazil
Catherine Salmon, University of Redlands, Redlands, California, USA
Emily Vogel, Northern Arizona University, Flagstaff, USA
Marina Butovskaya, National Research University Higher School of Economics, Moscow, Russia
ABSTRACT
Inclusive fitness theory postulates that preferentially provisioning offspring that most closely resemble the parents can indirectly increase their gene-copying success, with phenotypic resemblance between (allo)parent and offspring acting as an indicator of genetic similarity. According to this prediction, the amount of alloparental effort should correlate with parent-offspring resemblance cues, and this effect should be more pronounced for fathers and paternal kin, as paternity certainty is concerned predominantly with men. We tested these predictions and conducted an online survey in 2019 in Brazil (N = 605), Russia (N = 302), and the USA (N = 308).
By examining the relationship between parent-child resemblance and kin altruism, we have uncovered a widespread positive correlation between parent-child resemblance and the willingness of parents and grandparents to provide care. For example, the more a child resembles their father, the more parents and grandparents are likely to provide support. Similarly, the resemblance between a mother and child is positively associated with (allo)parental effort. At the same time, greater similarity between father and child can actually discourage matrilineal grandparents fr om providing childcare. In this respect, our data challenge the conventional viewpoint that paternity uncertainty and father-child similarity cues are the driving force behind paternal kin investments, but not maternal ones. Our results suggest that a more general kin recognition mechanism is at play, one that is shared by both matrilineal and patrilineal relatives in modern societies. One possible explanation is that in the context of the extended family, mothers and maternal relatives, as primary caregivers, may be particularly sensitive to key phenotypic traits of dependent children.
In the resulting model a sufficient cross-cultural difference emerges when examining the degree of assistance provided by different (allo)parents across the studied samples. For instance, American respondents highlighted a significant level of paternal involvement in childcare, while Russian respondents noted a high level of maternal kin assistance. In contrast, the Brazilian sample exhibited relatively low levels of kin involvement. The divergent paths of these countries raise important questions about the future of family structures and the role of kinship in shaping them. The highly urbanized Brazilian sample may provide valuable insights into possible future directions in family structure and the role of alloparental care within it. We propose that Brazil's high social integration and family member's incorporation into expanded social networks may contribute to the development of a communal model of breeding, marked by ultra-social or eusocial childcare practices. Will the Russian and American models of family follow the same ultra-social path as the Brazilian one? Or will Russian and US families maintain a distinct approach to childcare in a world influenced by globalization? We suppose that further field work on alloparental care in the Latin American region is essential to shed light on this important topic, and to uncover the answers to these intriguing questions.
Keywords: parent-child resemblance, grandparents, paternity certainty, kin-recognition, cooperative breeding.
1. EVOLUTION OF ALTRUISM
The concept of parental favoritism has long fascinated researchers and observers, who seek to understand why parents often show a strong preference for one child over others. While this phenomenon is often associated with evolutionary and genetic factors, it is essential to consider the broader context of altruism, which plays a crucial role in the development of social behaviors.
Peter Kropotkin is one such intellectual who was one of the first to draw attention to the importance of altruism in evolution, shedding light on the intricate social dynamics between individuals and groups. An anthropologist and philosopher, Kropotkin emphasized the importance of mutual aid within and between species in his works. In his book Mutual Aid: A Factor of Evolution (1902) Peter Kropotkin challenged the conventional view that survival is based on competition, arguing instead that cooperation and mutual aid have been essential for the development of societies and the emergence of altruism. Kropotkin drew inspiration fr om a rich tradition of Russian thought that emphasized community, mutual responsibility, and collective action, in contrast to the prevailing Western individualism and competition of the time, which was also widely rejected by the late nineteenth-century Russian intellectual circles (Gould 1988). Kropotkin's merit is that he succeeded in expressing these sentiments in the form of a coherent evolutionary theory and in presenting it to a broad Western audience.
But in the middle of the twentieth century, when the triumphant uni on of population genetics and Darwinism had conquered the minds of scientists, Kropotkin's concept of mutual aid had already been forgotten in the West. And only in the early 1960s, after the publication of the work of the English ornithologist Vero Copner Wynne-Edwards, Animal Dispersion (1962), did the topic of the evolution of mutual aid again become the subject of a heated debate. In this work, Wynne-Edwards noted that in natural environments untouched by man, unlimited population growth and the subsequent stage of complete depletion of the resource base are very rare: the normal evolutionary trend goes in the opposite direction, towards the creation and maintenance of a habitat at the highest achievable level. Like the Russian naturalists, he made his observations in a harsh climate zone – in Northern Canada, where Wynne-Edwards found that in seabird colonies less than half of the individuals begin to lay eggs each season. He suggested that the birds may deliberately abstain fr om reproduction in order to reduce intraspecific competition and increase their group's chances of success under conditions of resource scarcity. According to Wynne-Edwards, there is some homeostatic effect that maintains the number of breeding individuals and allows the population density to be balanced at an optimal level.
However, the main difficulty in explaining this evolutionary mechanism, capable of regulating the number of individuals for the benefit of the entire group and limiting their intraspecific aggressiveness in the struggle for limited resources, was that it conflicts with the classical concept of Darwinian fitness, which was based on the power of natural selection to maintain the spread of traits by increasing the number of descendants of the most adapted individuals. The difficulty of explaining the mechanism of intra-group selection proposed by Wynne-Edwards, as noted in Maynard Smith's 1964 work ‘Group Selection and Kin Selection’, also lies in the fact that in the presence of contacts between neighboring groups, ‘infection’ with the gene of antisocial behavior undoubtedly occurs, and the gene of asociality can quickly spread in any group of altruists. As a result of the presence of these contradictions, which were unresolvable at the time, the Theory of Group Selection did not find understanding among the evolutionists of the second half of the twentieth century.
A turning point in the study of the problems of altruism and the features of kin selection was made by William D. Hamilton. In 1964 in his famous paper ‘Genetic Evolution of Social Behavior’ (Hamilton 1964a, 1964b) he proposed the concept of inclusive fitness. Hamilton's work was built upon the idea that individuals can increase their reproductive success not only through the production of offspring, but also through the survival and reproduction of their relatives. Fr om his point of view, the main actors in evolution are not organisms, and certainly not groups or species, but genes, whose sole task is the maximum multiplication of their own copies. He presented a mathematical explanation of altruism as follows: an extreme degree of altruism, consisting in the sacrifice of one's own life for the good of others, can be supported by selection if this sacrifice can save more than two siblings, or more than four cousins. Self-sacrifice and altruism can also extend to more and more distant relatives, since common altruistic genes may be even more numerous in an extended social group, and even taking into account the distance of kinship, the ratio of their number to the coefficient of relatedness will have a positive effect on the maintenance of altruistic traits in a population. Some similar illustrations were given earlier by Haldane (1932, 1955), who, in fact, introduced the very concept of ‘altruistic behaviour’ into the thesaurus of evolutionary biology, pointing out that a behaviour such as an alarm call may harm the individual but benefit other members of the group. Haldane suggested two processes by which it might evolve. The first was the process of between-group selection, in which groups capable of achieving mutualistic goals eventually outcompete non-altruistic groups of egoists. The second model of the spread of altruistic traits, proposed by Haldane, was in many ways close to what we today know as kin selection theory. But it was in the hands of William Hamilton (1964a, 1964b) and his followers that the second one – the theory of kin selection, an evolutionary process which is used to be known as a key driver for the emergence of cooperative and altruistic behavior, – became a cornerstone of modern behavioral ecology (West, Griffin and Gardner 2007).
It should be noted that Hamilton (1975) upd ated his earlier theory to distinguish between kin selection theory and inclusive fitness theory. This distinction was based on whether or not the shared genes had been inherited by recent common descent. Hamilton explained that recent common descent had been no more than a simplifying assumption in his original kin selection model, stating that this had been done to render the mathematical model more manageable, but that this assumption was not a necessary one for his inclusive fitness theory. With the freeing of that theoretical constraint, inclusive fitness theory became virtually indistinguishable from the genetic similarity theory that was elaborated later (Rushton 1989). This is important because recent common descent is not the only way that the same genes can be shared between two or more individuals. Another generative mechanism is the production of offspring via positive assortative mating for genetic similarity. When that occurs, which is the norm in most modern human populations (Figueredo and Wolf 2009; Figueredo et al. 2015; Wolf and Figueredo 2011), then the coefficient of relatedness may be higher than the theoretical value of r=0.5 (under assumptions of panmixia, or random mating), depending on the degree of positive assortative mating.
Nowadays the evolution of altruism is a complex and ongoing topic in the field of evolutionary biology. Since the discovery of indirect fitness theory, experiments and modeling have made it possible to demonstrate the quantitative mechanisms of preservation and evolution of altruism in conditions unfavorable for the cooperating individual. Altruism and cooperation have been studied in various contexts, including public goods games, social dilemmas and economic interactions. Research has identified several triggers that facilitate cooperation among individuals such as direct and indirect reciprocity, when individuals reciprocate kindness or help with kindness in return (Nowak and Sigmund 1998), regular contact with others also can encourage cooperation (Falk and Fischbacher 2006) and imposing consequences for non-cooperation can help deter antisocial individuals from free-riding and encourage them to contribute to the collective effort (Barron 2024).
At the same time, debates continue among evolutionary theorists about levels of selection, human and animal social behavior, sexual selection, and related topics. Researchers such as Lee Cronk (Cronk 2019), Richard Wrangham (Wrangham 2021; Glowacki et al. 2020), Christopher Boehm (Boehm 2018) and Jon Haidt (Haidt 2007) contribute to the ongoing debate and investigations in the field of group selection, altruism, and cooperation. Their work explores the mechanisms and outcomes of evolutionary forces on complex behaviors, such as cooperation based on moral foundation, self-domestication and human social behavior. In recent years, the theory of multilevel selection has gained significant support, offering a potential explanation for human ultrasociality – the ability to cooperate in large groups of genetically unrelated individuals. This development has opened up new opportunities for interdisciplinary research, facilitating the convergence of history and biology, among other fields (Hertler et al. 2020). New approaches are also emerging, for example, Ohad Lewin-Epstein and Lilach Hadany (Lewin-Epstein and Hadany 2020) have proposed moving away from the individual and gene-centric picture of interactions and considering the microbes carried by the interacting individuals. The ongoing research in these fields by these and other scholars has helped to elucidate the mechanisms and outcomes of evolutionary forces on complex behaviors, including altruism and kin selection.
1.1 The Problem of Determining Relatedness
Hamilton's Rule of kin selection states that altruistic behavior for the benefit of unrelated individuals will only occur if the recipient of the benefit is not too distantly related, and the cost of helping is not too high. As a result, the ability to adequately determine kinship may become a key point in the manifestation of behavioral altruism, especially when the cooperative individual sacrifices significant efforts and resources, as is common, for example, in the context of kinship interactions that promote mutual help and care.
Due to its universal presence in human societies, kinship has traditionally been a core organizing principle in non-Western, non-urban societies that social anthropologists have focused on. It is not surprising, then, that many social anthropologists initially resisted the application of Hamilton's Rule to humans (Cronk 2019). Marshall Sahlins, a prominent anthropologist, was particularly critical of Hamilton's Rule, arguing that few humans could perform the complex mental calculations necessary to guide their behavior (Sahlins 1976). This skepticism was later labeled ‘Sahlins' Fallacy’ by evolutionary biologists (Dawkins 1979: 291–292), who view it as a cautionary tale in evolutionary psychology.
The underlying problem lies in the fundamentally different assumptions that social scientists and evolutionary biologists make about human behavior. Social scientists like Sahlins assume that behavior is the result of conscious deliberation shaped by culture, whereas evolutionary biologists recognize that many evolved mechanisms operate below the threshold of conscious awareness. The hypothetical mechanism for the evolution of altruistic behavior toward individuals with phenotype tags that are sufficiently similar to the donor's phenotype was elaborated upon by Dawkins. In 1982, in his work on the extended phenotype question, Richard Dawkins proposed a specific innate mechanism of kin recognition that might operate between genealogical relatives. He gave it an olfactory name – the so-called armpit effect (Dawkins 1982). Dawkins supposed that organisms might follow the rule ‘be nice to kin who smell more similar to you’. This mechanism potentially facilitates the identification of a degree of genetic relatedness via phenotype matching, which involves individuals' ability to recognize a particular phenotypic marker carried by both the individual and its partner and to behave nepotistically on the basis of their phenotypic similarity (Dawkins 1982; Hamilton 1964a, 1964b; Mateo and Johnston 2000; Rousset and Roze 2007). Later, the recognition mechanism that discriminates between different recipients of altruistic help on the basis of such a phenotypic similarity was positively confirmed (Traulsen and Nowak 2007; Bavik et al. 2022).
In humans, the recognition between close and distant kin, as well as non-kin also employs an elaborate kinship family system (Jones 2004; Hamilton 1964a, 1964b; Hames 2015). However, there are still situations wh ere an individual's relatedness to others can be ambiguous. Due to internal fertilization in humans, males, and paternal relatives may have greater uncertainty about their genetic relatedness to offspring than mothers and maternal kinships. In evolutionary psychology, the issue of unclear biological relatedness to the offspring for fathers and paternal kin is known as the theoretical concept of paternal uncertainty. Fitness losses for men who allocate their reproductive effort in parental care for unrelated children can be considerable, and males are expected to invest less in children whose relations are putative (Apicella and Marlowe 2004).
The issue of intrafamilial altruism and relatedness uncertainty also relates to patrilineal kin effort, as the paternity threshold model formalizes the role of paternity uncertainty as an adaptive explanation for biases in alloparental effort (Kurland 1979; Perry and Daly 2017). A number of studies have found that matrilineal grandparents are more inclined to invest in grandchildren than patrilineal grandparents (Perry and Daly 2017; Sear et al. 2000; Voland and Beise 2002).
Reasonably, in the past there were no explicit ways for fathers and patrilineal relatives to verify biological relatedness to a child. Therefore, a process of self-referent phenotype matching and recognition of signs of physical and personality cues of kin resemblances could help the kin to determine the presence of biological relatedness and facilitate alloparental decision making about care. A rapidly growing body of research has demonstrated the human ability to match the phenotypic tags based on visual images, and to fairly accurately detect the actual genetic relatedness between different faces (Alvergne et al. 2009; Brédart and French 1999; Fasolt 2021; Woo 2021), even for faces of different races (Alvergne et al. 2009; Woo 2021). For instance, several experiments have shown that people's judgments on the facial similarity of child faces correlate with their actual relatedness (Maloney and Dal Martello 2006; DeBruine et al. 2009). In experiments appealing to the actual or digitally modified images have shown that similarity cues generally lead to more positive attitudes towards children who resemble adult subjects, and even guide fathers' decision making about parental effort (Platek et al. 2004; Volk and Quinsey 2002).
However, paternity uncertainty is not the only adaptive function for determining one's degree of genetic relatedness to a given potential recipient of parenting or alloparenting. Higher coefficients of relatedness are predicted to produce higher levels of altruism, so greater degrees of positive assortative mating should indirectly sel ect for higher levels of child-directed parenting and alloparenting, and this logic should apply to the mother and matrilineal relatives as well as to the father and patrilineal relatives, even in the absence of maternity uncertainty. Thus, monitoring the degree of relatedness to a given child by the mother and matrilineal relatives for the purposes of optimal parenting and alloparenting would be theoretically expected when the degree of positive assortative mating between the parents of the children may be variable (Figueredo and Wolf 2009; Figueredo et al. 2015; Wolf and Figueredo 2011).
1.2 Current Research Design
The goal of this paper was to test whether the amount of alloparental effort is correlated with parent-offspring resemblance cues, and whether this effect is more pronounced for fathers and paternal kin, compared to mothers and maternal kin in a cross-cultural perspective.
We hypothesized that Father-Child resemblance traits in a child would enhance Paternal Effort and Patrilineal kin care in all studied cultures. In contrast, Mother-Child resemblance should demonstrate a relatively modest impact on maternal and alloparental help.
It is hypothesized that culturally specific kinship structures may also play an important role in caregiving, as the availability of matrilineal and patrilineal kin may vary considerably. Here we implement a cross-cultural approach to test the effect of parent-child similarities in three post-industrial societies: Brazil, Russia, and the USA. These three countries are comparable in a number of important aspects, including their population and territorial size, urbanization rates and levels of integration into the global economy (www.cia.gov/the-world-factbook/countries/). All the three countries have a solid European cultural heritage, namely their main ethnic groups profess Christianity, implying that monogamy (as well as low extra-pair paternity rates) has been the only legitimate form of marriage there for centuries.
2. METHODS
2.1 Samples
Brazil. Data from Brazil were collected online from college students at two different public universities: the University of São Paulo, in the state of São Paulo and the Federal University of Espírito Santo, in the state of Espírito Santo, both in southeastern Brazil.
Russia. Data for the Russian sample were collected online. Participants were recruited from students at the Moscow Institute of Physics and Technology and also through a series of advertisements on the Russian segment of the Internet mostly through social media facilities.
United States. Data for the United States were collected online in two states: Arizona and California. The Arizona sample was collected from college students at the University of Arizona in Tucson, AZ. The California sample was collected from college students at the University of Redlands in Redlands, CA. Participants were compensated with course credit.
Table 1
Demographic information on respondents
from Brazil, Russia, USA
| Brazil | Russia | USA |
N | 603 | 301 | 308 |
Age (M) | 25.5 | 29.1 | 19.6 |
Age (SD) | 9.28 | 9.82 | 2.33 |
Age Range | 16–67 | 17–67 | 18–38 |
Female n | 468 (78 %) | 214 (71 %) | 231 (74 %) |
Male n | 133 (22 %) | 88 (29 %) | 81 (26 %) |
2.2 Measures
Parent-Child Resemblance Scales. To assess the degree of similarity between a child and a parent, our team utilized five-item subscales measuring self-perceived parent-child similarity. Participants rated the degree of resemblance to each of their parents (e.g., ‘Overall resemblance’, ‘Hair color and structure resemblance’) on a 6-point scale (wh ere 0 = not very similar and 5 = very similar). In order to assess the fit between the observed data and theoretically grounded constraints of Parent-Child Resemblance, all items within this scale underwent a CFA procedure (Appendix A).
Parental Effort Subscales. The Early Environment Questionnaire was used to represent the latent constructs of maternal and paternal direct care using a 6-point scale. The final subscale included items such as: ‘I felt that my mother was emotionally supportive’; ‘My father usually helped me solve problems’. To assess the fit between the observed data and the theoretically grounded constructs of Maternal and Paternal Effort, all items were subjected to CFA procedure (Appendix A).
Grandparental Assistance. To assess the level of grandparental Assistance our team utilized three-item subscales. Sample items include questions like: ‘During my childhood, my maternal grandparents lived: with us; separately, but in the same city; in another city, but no more than three hours from us; in another city, more than three hours away from us; in another country; I mostly lived with my grandparents and not my parents?’; ‘How often did you spend your school holidays with your paternal grandparents?’ Participants used a 5-point scale (0 never; 4 almost always) to describe the frequency of each behavior during the childhood. To assess the fit between the observed data and the theoretically grounded constricts of Maternal and Paternal Grandparental Assistance all items underwent a CFA procedure (Appendix A).
Bayesian Multilevel Modeling (BMLM) was implemented to analyze the (allo)parental effort given by four family agents to a child based on the degree of resemblance the child has with its mother and father. In this model, both the IDs of the participants and the national group to which they belong were included as grouping factors in order to account for inter-individual correlations within the samples studied on individuals' responses.
3. RESULTS AND DISCUSSION
3.1 Understanding the Role of Father-Offspring Resemblance in Kin Care
The summary output of the fitted BMLM model reveals that investments in childcare by both parents and grandparents appear to be positively mediated by the physical similarity between a child and a parent (similarity to a son or daughter in the case of grandparents).
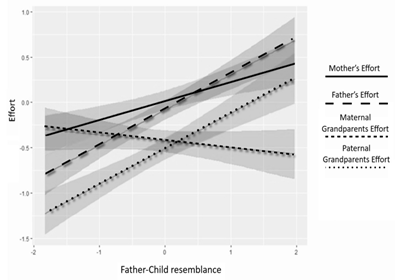
Fig. 1. Visualization of the Father-child resemblance factor
on the level of effort of four alloparents
For instance, father-child resemblance demonstrated a positive regression coefficient and reliable confidence intervals in the BMLM model, which implies that paternal effort, which is a baseline in our model, is significantly higher for a child resembling the father (b = .39; CI [.28, .5]). However, it is worth noting that father-child resemblance has a significant negative impact on matrilineal kin care, specifically on the maternal grandparent's assistance (b = –.46; CI = [–.33; –.03]) and the mother care (b = –.18; CI= [–.33; –.03]).
3.2 Understanding the Role of Mother-Offspring Resemblance in Kin Care
The summary output displays a positive regression coefficient (b = .11) for mother-child resemblance, although the 95 % credibility interval (CI = [–.01; .23]) is relatively wide. The wide credibility interval in our data suggests that there is not enough evidence to reasonably estimate the impact of mother-child resemblance as a significant positive predictor influencing alloparent's attitudes. However, mothers themselves do respond to phenotypic cues of similarity between themselves and their children. And mothers significantly increase their efforts towards children who resemble them (b = .21, 95; CI = [.05; .37]).
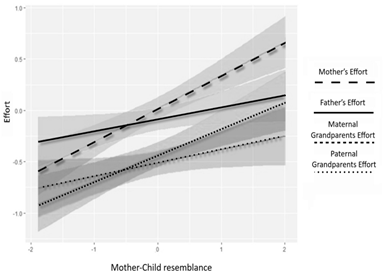
Fig. 2. Visualization of the mother-child resemblance factor
on the level of effort of four alloparents
3.3 The Combined Effect of Culture and Alloparent
An interaction between the categorical variables is shown in Figure 3, which shows the combined effect of the two factors: country and alloparent. According to our analysis, the overall paternal effort was higher in the United States compared to Russia and Brazil (b = .28, 95 %; CI =
= [.09; .47]). This suggests that, holding other predictors constant, children in the USA receive more kin assistance than in the other two countries. In contrast, Russia showed a notably higher availability of maternal helpers, who invested significantly more in their grandchildren compared to the other studied countries. Furthermore, maternal kin help in the Russian sample stands out as the highest of all the types of alloparental effort we examined (b = .71, 95 %; CI = [.41; 1.01]).
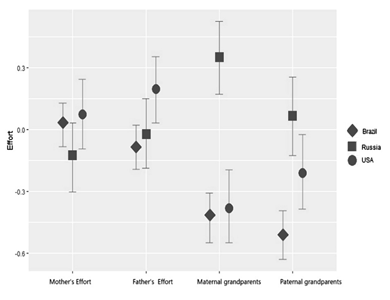
Fig. 3. Visualization of the combined marginal effects
of four alloparental effort across three cultures
4. CONCLUSIONS AND POLICY IMPLICATIONS
4.1 Alloparents Care and Parent-Offspring Resemblance
While children share half of their genetic information with each parent and only about a quarter with their grandparents, both paternal and maternal family members invest in ensuring the well-being of their dependent children. However, confidence in the genetic relationship with their grandchildren varies among family members, including grandmothers and grandfathers from both sides of the family. Theoretical predictions suggest that in humans, maternal relatives and mothers are more certain about their genetic relationship with offspring and therefore more interested in caring for them. In contrast, paternal grandparents may have less confidence in genetic relatedness due to uncertainty about paternity. Given these factors, paternal grandparents may be more attuned to the phenotypic similarity between their son and his children, and adjust their investment in grandchildren according to signs of similarity to their sons.
In this paper, we test this hypothesis using cross-cultural questionnaire data collected in 2019 in three countries: Russia, the USA, and Brazil. The collected data were analyzed using Bayesian Multilevel Modeling (BMLM) to predict overall (allo) parental effort based on the self-reported similarity levels between the participants and their mother and father.
BMLM analyses have shown that investments in childcare by both parents and grandparents appear to be positively mediated by the physical similarity between a child and a parent (similarity to a son or daughter in the case of grandparents). The graphical visualization of the results demonstrated the generally positive impact of the similarity factors on the effort of each family member, but not all regressions were statistically significant.
On the one hand, the association between care from patrilineal relatives and father-child similarity was found to be significant and positive (Figure 1). Оn the other hand, we also observed a significant positive correlation between mother-child similarity and the effort put forth by mothers and maternal relatives in childcare (Figure 2). Additionally, maternal relatives showed negative reactions to the child's physical appearance, leading to a significant decrease in their effort when prominent signs of resemblance between the child and the father were present (Figure 1). In contrast, as shown in Figure 2, other relatives, including patrilineal kin, generally viewed the mother-child resemblance positively.
One possible explanation for the positive response of matrilineal relatives to similarity cues and the increasing hostility to alien traits is an adaptive response to cues of genetic relatedness in different social contexts (DeBruine 2004). Based on the premise that in most mammals, including humans, young are raised in family groups with substantial variation in genetic relatedness among kin family members, Mark E. Hauber and Paul W. Sherman (2001) developed the concept of adaptive innate neural recognition mechanisms to deal with physical similarity clues. In a human family in the prosocial context of cooperative breeding, offspring develop in a family of mixed relatedness, such as through multiple paternity or maternity, or among unrelated conspecifics. Individuals reared together may have different sires, so that full and half siblings grow up side-by-side.
However, the phenotypic sensitivity of females cannot be fully explained by the lack of biological relatedness that may exist in extended family contexts. Mixed relatedness is normative among full siblings and their offspring due to genetic recombination processes during the meiotic phase of reproduction, which result in a novel se t of genetic information in each half of the ancestral genetic information passed from parents to their common children. Therefore, phenotypic similarity does not represent a completely irrelevant kinship cue for maternal relatives because the combination of positive assortative mating (which is prevalent in humans for most permanent and stable traits) and random genetic recombination may result in maternal relatives sharing more genetic similarity with one of their offspring than another (Rushton 1988; Nivard et al. 2024; Robinson et al. 2017).
One of the main conclusions that could be drawn from this article is that female effort depends on the perceived mother-child similarity factor, and that finding is most likely consistent with the evolutionary framework of the cooperative breeding model as a pervasive reproductive adaptation in humans, as explicitly delineated in the work of Sara Hrdy (1999, 2007) and Karen Kramer (2005, 2010).
4.2 Childcare through the Lens of Cross-Cultural Analysis
The sociological approach to cultural dimensions, as outlined by Geert Hofstede, classifies Russia and Brazil as collectivistic cultures, emphasizing the prioritization of the group's needs over individual needs (Hofstede 2011). On the other hand, the United States is an individualistic culture, wh ere individuals are expected to prioritize their own needs and interests over those of the group. The analysis of alloparental effort in the studied countries partially supported these predictions: American participants demonstrated higher levels of self-efficacy in caregiving compared to those from other countries, indicating that they felt more confident in their ability to care for their children and less reliant on alloparents. American fathers have relatively high involvement in childcare, compared to participants from other countries. This suggests that cultural norms in the United States may be supportive of greater paternal effort, which can lead to better outcomes for children. It is also evident that Americans are more likely to identify family norms as more gender neutral, with fathers more involved in childcare.
In contrast to other countries, Russia has demonstrated a remarkable prevalence of matrilineal helpers who heavily invest in their grandchildren. Our data reveal that the continuation of family traditions in Russia is largely maintained by maternal kin, with a significant presence of maternal grandparents in daily care and other family matters. This shift towards a matrilineal family structure in modern Russia reflects a profound cultural transformation: the evidence suggests that Russia has moved away from a patrilocal, patriarchal society, which is generally considered normative for Russian society (see, e.g., Gapirzhanovna 2023), and towards a more egalitarian family structure in which maternal relatives play a more prominent role (Semenova and Butovskaya 2022).
In stark contrast to Russia, Brazilian grandparents show a marked disinterest in providing childcare assistance. This is particularly evident among paternal grandparents, who show little enthusiasm for childrearing. While some researchers argue that racial and ethnic differences may influence grandparental support in different cultures (Livingston and Parker 2010), we suggest that the high rates of urbanization in Brazil may be a contributing factor. Potentially, the significant migration processes and rapid urbanization of Brazilian cities in recent decades (Aguayo-Tellez et al. 2010) may have led to a shift away from traditional kinship support.
In this respect, the decline in familial support in Brazil may be linked to several factors. Firstly, it is possible that many families lack the financial means to maintain frequent contact with their relatives living in a distant city. Secondly, it may be a consequence of the inadequate development of national transport networks (Ferreyra and Roberts 2018). Finally, it may reflect cultural changes in the family structure, influenced by the rapid population growth in Brazilian megacities, leading to increased involvement of genetically unrelated community members in caregiving practices. Ethnographic data point to the high importance of social integration for members of Brazilian society, who are integrated into expanded social networks under the conditions of growing urban agglomerations (Klippel 2018). The American-born Brazilian anthropologist Claudia Fonseca estimates that in poor neighborhoods about half of the women give their children to other families or to state-run shelters, either on a short or long-term basis. In working-class areas, wealthier families are more likely to take in other people's children. A 1986 study by Claudia Fonseca in Porto Alegre, the largest city in southern Brazil, found that of the 120 households she surveyed, about one hundred respondents had lived with adoptive mothers, godparents, or grandmothers, often called mothers de criação (Fonseca 1995). This arrangement allowed for childcare assistance to be provided by the social group, rather than solely by genetic relatives. Social care for children in the context of shared parenthood can be seen as an example of eusocial characteristics in contemporary Brazilian society (Cassar et al. 2023), potentially facilitating adaptability to local childcare needs and the exchange of support and resources between unrelated individuals (Fonseca 2003).
4.3 Policy Implications
Our study has shed light on the intricate interplay of cultural and biosocial factors that shape family dynamics in three different regions: Russia, Brazil, and the United States. In this complex tapestry, we discovered both universal threads of parental support and unique patterns characteristic of each region. For instance, in the United States, the prevalence of an individualistic culture and strong paternal involvement in childcare should be celebrated as a positive trend. To support these emerging norms, policymakers can encourage flexible work arrangements and family-friendly policies that recognize the importance of paternal involvement in childcare and develop programs and initiatives that promote intergenerational family relationships and reinforce a sense of community responsibility. In Russia, a unique reliance on matrilineal kin support has emerged, highlighting the vital role that extended families play in shaping modern family dynamics in this region. Our research suggests that strengthening traditional family values and a sense of parental responsibility may be crucial for Russia. By embracing and supporting these values, we can create a stronger foundation for families and promote a culture of mutual care and support.
For Brazil, the societal shift towards more informal, collective forms of childcare offers an opportunity to strengthen social ties and create more resilient communities. By encouraging involvement of unrelated individuals in caregiving, Brazil can harness the flexibility and resourcefulness of its growing urban agglomerations. Brazil's high levels of social integration and inclusion in extended social networks may encourage the development of a more communal approach to childcare, resembling ultrasocial or eusocial caregiving practices.
The divergent paths of these countries raise fundamental questions about the future of family structures and the role that kinship plays in shaping them. Will the United States continue to emphasize individualized forms of childcare, in which fathers play a more prominent role? Will the role of grandmothers remain as important in Russia? Or will the increasingly urbanized Brazilian model become a harbinger of a new, more communal approach to childcare? In this sense, how prophetic were the ideas of Peter Kropotkin, who noticed that social organisms such as insects and some species of mammals form a complex structural organization with a clear division of labor and cooperation between individuals, and in which he saw parallels with human societies, wh ere people often pool their efforts together to achieve a common goal.
As we ponder these questions, we are reminded of the vast cultural and economic disparities that shape family dynamics around the globe. In a world marked by increasing global connectivity and social integration, it is likely that family structures will continue to evolve, integrate and adapt. The fascinating trajectories of the United States, Russia, and Brazil provide a unique opportunity to explore the complex interplay between culture, biology, and society in shaping family structures. Further fieldwork on alloparental care in the Latin American region is essential to shed light on this important topic, and to uncover the answers to these intriguing questions.
5.0 DATA AVALABILITY
FUNDING
This research has been supported by the Russian Science Foundation (project № 24-28-01639, https://rscf.ru/project/24-28-01639/). (OS)
REFERENCES
Aguayo-Tellez, E., Muendler, M. A., and Poole, J. P. 2010. Globalization and Formal Sector Migration in Brazil. World Development 38 (6): 840–856.
Alvergne, A., Oda, R., Faurie, C., Matsumoto-Oda, A., Durand, V., and Raymond, M. 2009. Cross-Cultural Perceptions of Facial Resemblance between Kin. Journal of Vision 9 (6): 23–23.
Apicella, C. L., and Marlowe, F. W. 2004. Perceived Mate Fidelity and Paternal Resemblance Predict Men's Investment in Children. Evolution and Human Behavior 25 (6): 371–378.
Barron, E. N. 2024. Game Theory: An Introduction. John Wiley and Sons.
Bavik, L., Mehta, R., and Weissman, D. 2022. Robust Altruism by Similarity in Multifaceted Phenotypes. APS March Meeting Abstracts 2022 (1): 5–12.
Brédart, S., and French, R. M. 1999. Do Babies Resemble Their Fathers more than Their Mothers? A Failure to Replicate Christenfeld and Hill (1995). Evolution and Human Behavior 20 (2): 129–135.
Boehm, C. 2018. Collective Intentionality: A Basic and Early Component of Moral Evolution. Philosophical Psychology 31 (5): 680–702.
Cassar, A., Cristia, A., Grosjean, P. A., and Walker, S. 2023. It Makes a Village: Allomaternal Care and Prosociality. SSRN eLibrary, 19. URL: http:// research.economics.unsw.edu.au/RePEc/papers/2022-06.pdf.
Cronk, L. 2019. That Complex Whole: Culture and the Evolution of Human Behavior. Routledge.
Dawkins, R. 1982. The Extended Phenotype: The Gene as the Unit of Selec-tion. Oxford University Press.
Dawkins, R. 1979. The Selfish Gene. New York: Oxford University Press.
DeBruine, L. M. 2004. Resemblance to Self Increases the Appeal of Child Faces to both Men and Women. Evolution and Human Behavior 25 (3): 142–154.
DeBruine, L. M., Smith, F. G., Jones, B. C., Roberts, S. C., Petrie, M., and Spector, T. D. 2009. Kin Recognition Signals in Adult Faces. Vision Research 49 (1): 38–43.
Falk, A., and Fischbacher, U. 2006. A Theory of Reciprocity. Games and Eco-nomic Behavior 54 (2): 293–315.
Fasolt, V. 2021. Third-Party Kinship Recognition. Doctoral dissertation. Uni-versity of Glasgow.
Ferreyra, M. M., and Roberts, M. (Eds.) 2018. Raising the Bar for Productive Cities in Latin America and the Caribbean. World Bank Publications.
Figueredo, A. J., and Wolf, P. S. A. 2009. Assortative Pairing and Life History Strategy: A Cross-Cultural Study. Human Nature 20: 317–330.
Figueredo, A. J., Wolf, P. S. A., Olderbak, S. G., Sefcek, J. A., Frías-Arme-nta, M., Vargas-Porras, C., and Egan, V. 2015. Positive Assortative Pairing in Social and Romantic Partners: A Cross-Cultural Observational Field Study of Naturally Occurring Pairs. Personality and Individual Differences 84: 30–35.
Fonseca, C. 1995. Caminhos da adoção. 3a edição. São Paulo, Cortez.
Fonseca, C. 2003. Patterns of Shared Parenthood among the Brazilian Poor. Social Text 74: 111–127.
Gapirzhanovna, T. D. 2023. Russian Patriarchal Family in its Structure and Composition. Journal of Innovation, Creativity and Art: 75–78.
Glowacki, L., Wilson, M. L., and Wrangham, R. W. 2020. The Evolutionary Anthropology of War. Journal of Economic Behavior & Organization 178: 963–982.
Gould, S. J. 1988. Kropotkin was no Crackpot. Natural History 97 (7): 12–21.
Haidt, J. 2007. The New Synthesis in Moral Psychology. Science 316 (5827): 998–1002. doi:10.1126/science.1137651.
Haldane, J. B. S. 1932. The Causes of Evolution. London: Longmans Green.
Haldane, J. B. S. 1955. Population Genetics. Penguin New Biol. 18: 34–51.
Hames, R. 2015. Kin Selection. Anthropology Faculty Publications: 505–523.
Hamilton, W. D. 1964a. The Genetical Evolution of Social Behaviour. I. Journal of Theoretical Biology 7: 1–16.
Hamilton, W. D. 1964b. The Genetical Evolution of Social Behaviour. II. Journal of Theoretical Biology 7 (1): 17–52.
Hamilton, W. D. 1975. Innate Social Aptitudes of Man: An Approach from Evolutionary Genetics. Biosocial Anthropology 133: 115–132.
Hauber, M. E., and Sherman, P. W. 2001. Self-Referent Phenotype Matching: Theoretical Considerations and Empirical Evidence. Trends in Neuro-sciences 24 (10): 609–616.
Hertler, S. C., Figueredo, A. J., and Peñaherrera-Aguirre, M. 2020. Multilevel Selection: Theoretical Foundations, Historical Examples, and Empirical Evidence. Springer Nature.
Hofstede, G. 2011. Dimensionalizing Cultures: The Hofstede Model in Context. Online Readings in Psychology and Culture 2 (1): 8.
Hrdy, S. B. 1999. Mother Nature: A History of Mothers, Infants and Natural Selection. New York, NY: Pantheon.
Hrdy, S. B. 2007. Evolutionary Context of Human Development: The Coope-rative Breeding Model. In Salmon, C. A., and Shackelford, T. K. (eds.), Family Relationships: An Evolutionary Perspective (pp. 39–68). Oxford University Press.
Jones, D. 2004. The Universal Psychology of Kinship: Evidence from Lan-guage. Trends in Ecology & Evolution 8 (5): 211–215.
Klippel, A. P. Z. 2018. Paternidade socioafetiva: a família e sua evolução histórico cultural. Diss. Palhoça.
Kramer, K. L. 2005. Children's Help and the Pace of Reproduction: Coope-rative Breeding in Humans. Evolutionary Anthropology: Issues, News, and Reviews 14 (6): 224–237.
Kramer, K. L. 2010. Cooperative Breeding and its Significance to the Demo-graphic Success of Humans. Annual Review of Anthropology 39: 417–436.
Kropotkin P. 1902. Mutual Aid: Factor of Evolution. N. Y.: McClure, Phillips.
Livingston, G., and Parker, K. 2010. Since the Start of the Great Recession, More Children Raised by Grandparents. Pew Research Center, Septem-ber 9. URL: https://www.pewresearch.org/social-trends/2010/09/09/since-the-start-of-the-great-recession-more-chi....
Kurland, J. A. 1979. Paternity, Mother's Brother, and Human Sociality. Evo-lutionary Biology and Human Social Behavior: 145–180.
Lewin-Epstein, O., and Hadany, L. 2020. Host–Microbiome Coevolution can Promote Cooperation in a Rock–Paper–Scissors Dynamics. Proceedings of the Royal Society B, 287 (1920): 20192754.
Maloney, L. T., and Dal Martello, M. F. 2006. Kin Recognition and the Per-ceived Facial Similarity of Children. Journal of Vision 6 (10): 1047–1056.
Mateo, J. M., and Johnston, R. E. 2000. Kin Recognition and the ‘Armpit Effect’: Evidence of Self–Referent Phenotype Matching. Proceedings of the Royal Society of London. Series B: Biological Sciences, 267 (1444): 695–700.
Maynard Smith, J. 1964. Group Selection and Kin Selection. Nature 201 (4924): 1145–1147.
Nivard, M. G., Belsky, D. W., Harden, K. P., Baier, T., Andreassen, O. A., Ystrøm, E., ... and Lyngstad, T. H. 2024. More than Nature and Nurture, Indirect Genetic Effects on Children's Academic Achievement are Conse-quences of Dynastic Social Processes. Nature Human Behaviour 8 (4): 771–778. doi: 10.1038/s41562-023-01796-2.
Nowak, M. A., and Sigmund, K. 1998. The Dynamics of Indirect Reciprocity. Journal of Theoretical Biology 194 (4): 561–574.
Perry, G., and Daly, M. 2017. A Model Explaining the Matrilateral Bias in Alloparental Investment. Proceedings of the National Academy of Sciences 114 (35): 9290–9295.
Platek, S. M., Raines, D. M., Gallup Jr, G. G., Mohamed, F. B., Thomson, J. W., Myers, T. E., ... and Arigo, D. R. 2004. Reactions to Children's Faces: Males are More Affected by Resemblance than Females are, and so are Their Brains. Evolution and Human Behavior 25 (6): 394–405.
Robinson, M. R., Kleinman, A., Graff, M., Vinkhuyzen, A. A., Couper, D., Miller, M. B., ... and Visscher, P. M. 2017. Genetic Evidence of Assortative Mating in Humans. Nature Human Behaviour 1 (1): 0016. URL: https:// www.nature.com/articles/s41562-016-0016.
Rousset, F., and Roze, D. 2007. Constraints on the Origin and Maintenance of Genetic Kin Recognition. Evolution: International Journal of Organic Evo-lution 61 (10): 2320–2330.
Rushton, J. P. 1988. Genetic Similarity, Mate Choice, and Fecundity in Hu-mans. Ethology and Sociobiology 9: 329–335.
Rushton, J. P. 1989. Genetic Similarity, Human Altruism, and Group Selection. Behavioral and Brain Sciences 12: 503–559.
Sahlins, M. 1976. The Use and Abuse of Biology: An Anthropological Critique of Sociobiology. University of Michigan Press.
Sear, R., Mace, R., and McGregor, I. A. 2000. Maternal Grandmothers Improve Nutritional Status and Survival of Children in Rural Gambia. Proceedings of the Royal Society of London. Series B: Biological Sciences, 267 (1453): 1641–1647.
Semenova, O. V., Butovskaya, M. L. 2022. Matrilateral Kinship Investment in Grandchildren in a Modern Russian Family. Discrimination of Female Offspring by Grandparents. Sibirskie istoricheskie issledovaniya 1: 138–155. Original in Russian (Семенова, О. В., Бутовская, М. Л. Матри-латеральный родственный вклад во внуков в современной россий-ской семье. Дискриминация потомков женского пола со стороны ба-бушек и дедушек. Сибирские исторические исследования 1: 138–155).
Traulsen, A., and Nowak, M. A. 2007. Chromodynamics of Cooperation in Finite Populations. PLoS One 2 (3): e270.
Voland, E., and Beise, J. 2002. Opposite Effects of Maternal and Paternal Grandmothers on Infant Survival in Historical Krummhörn. Behavioral Ecology and Sociobiology 52 (6): 435–443.
Volk, A., and Quinsey, V. L. 2002. The Influence of Infant Facial Cues on Adoption Preferences. Human Nature 13 (4): 437–455.
West, S. A., Griffin, A. S., and Gardner, A. 2007. Social Semantics: Altruism, Cooperation, Mutualism, Strong Reciprocity and Group Selection. Jour-nal of Evolutionary Biology 20 (2): 415–432.
Wolf, P. S. A., and Figueredo, A. J. 2011. Fecundity, Offspring Longevity, and Assortative Mating: Parametric Tradeoffs in Sexual and Life History Strat-egy. Biodemography and Social Biology 52 (2): 171–183.
Woo, P. J. 2021. Face Processing in a Multiracial Environment: Differential Experience in Face Recognition, Face Categorisation and Kinship Detec-tion. Doctoral dissertation. Université Grenoble Alpes.
Wrangham, R. W. 2021. Targeted Conspiratorial Killing, Human Self-Domes-tication and the Evolution of Groupishness. Evolutionary Human Sciences 3: e26.
Wynne-Edwards, V. C. 1963. Intergroup Selection in the Evolution of Social Systems. Nature 200: 623–626. https://doi.org/10.1038/200623a0.
Wynne-Edwards, V. C., and Wynne-Edwards, V. C. 1962. Animal Dispersion in Relation to Social Behaviour. Edinburgh and London: Oliver and Boyd.
Appendix
Appendix A
Latent constricts and confirmatory factor analysis
Grandparents Assistance. Manifest variables were used in the hypothe-sized CFA model of the association between paternal and maternal kin. The tests of the convergent validities (a major component of a CFA) of the two latent constructs produced from the observed 6 variables were psychomet-rically acceptable. CFA parameters: Comparative Fit Index (CFI) = .85; Tucker-Lewis Index (TLI) = .72; Test statistic = 153.944; df = 8; p-value (Chi-square) < .001; Standardized Root Mean Square Residual (SRMR) = = .079. All the pathways showed P(> |z|) < .001, and regression Paternal Grandparents → Maternal Grandparents shows P(> |z|) = .003, with B = .097).
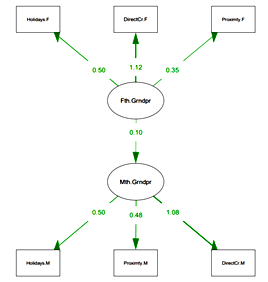
Fig. 1. CFA Grandparents Assistance model for latent variables results:
Maternal Grandparent Assistance
and Paternal Grandparent Assistance (two domains)
Notes: Fth.Grndpr represents the latent construct of paternal grandparent availability; Holiday.F – How often did you spend your school holiday with your grandparents on your father's side?; Proximity.F – ‘During my childhood my grandparents on my father's side lived’; DirectCr.F – ‘How often did your grandparents on your father's side babysit with you when you were ill?’; Mth. Grndpr represents the latent construct of maternal grandparent availability; Holiday.M – ‘How often did you spend your school holiday with your grandparents on your mother's side?’;Proximity.M – ‘During my childhood my grandparents on my mother's side lived’; DirectCr.M – ‘How often did your grandparents on your mother's side babysit with you when you were ill?’
Parent-Child Resemblance Scale. 10 items were used as observed variables in confirmatory factor analyses to depict the pattern of observed variables for higher ordered latent constructs: ‘Father-Child resemblance’ and ‘Mother-Child resemblance’ (Figure 2). Model parameters: Comparative Fit Index (CFI) =.84; Tucker-Lewis Index (TLI) = .79; Test statistic = 586.219; df = 34; p-value (Chi-square) < .001; Standardized Root Mean Square Residual (SRMR) = .085. All the pathways showed significant p(> |z|) < .001, except correlation Mother Resemblance ↔ Father Resemblance, p(> |z|) = .193; B = .047).
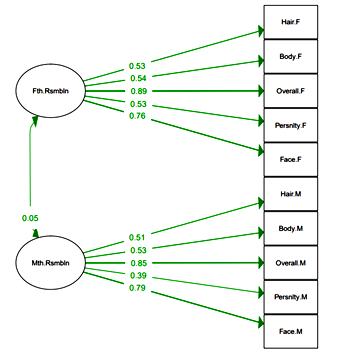
Fig. 2. Confirmatory factor analyses (CFA) for Latent variables results:
Parent-child resemblance constituting two domains:
Father-child resemblance (upper construct)
and Mother -child resemblance (lower construct)
Notes: Model items include: Fth.Rsmbln – father-child resemblance; Hair.F – Hair color and structure resemblance with father; Body.F – Body type resemblance with father; Overall.F – Overall resemblance with a father; Persnlty.F – Personality resemblance with a father; Mth.Rsmbln – mother-child resemblance; Hair.M – Hair color and structure resemblance with mother; Body.M – Body type resemblance with mother; Overall.M – Overall resemblance with mother; Persnlty.M – Personality resemblance with mother.
Parental Efforts Subscales. 8 items were used as observed variables in a CFA procedure to describe the pattern of observed variables for higher-ordered latent constructs: ‘Father Effort’, ‘Mother Effort’. The results of the CFA procedure are represented in Figure 3.
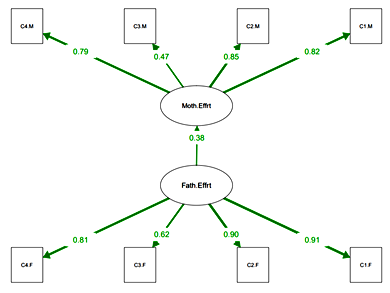
Fig. 3. SEM Parental Effort
Notes: Model items include: Mother helped solve problems (C1.M), maternal emotional support (C2.M), mother allowed learn from mistake (C3.M), amount of care and love that mother gave (C4.M), paternal emotional support (C1.F), father helped solve problems (C2.F), father allowed learn from mistake (C3.F), amount of care and love that father gave (C4.F).